Kromoszómák
Definíció - mi a kromoszóma?
A sejt genetikai felépítését DNS (dezoxiribonukleinsav) és bázisai (adenin, timin, guanin és citozin) formájában tárolják. Minden eukarióta sejtben (állatok, növények, gombák) ez a sejtmagban kromoszómák formájában van jelen. A kromoszóma egyetlen, koherens DNS-molekulából áll, amely bizonyos fehérjékhez kapcsolódik.
A kromoszóma név a görögből származik, és nagyjából lefordítható "színes testnek". Ez a név abból a tényből származik, hogy a citológia történetének nagyon korai szakaszában (1888) a tudósoknak speciális alapfestékekkel sikerült őket megfesteniük és fénymikroszkóppal azonosítaniuk. Ezek azonban a sejtciklus egy bizonyos pontján, a mitózisban (csírasejtekben meiózisban) láthatók igazán, amikor a kromoszóma különösen sűrű (kondenzált).

Hogyan épülnek fel a kromoszómák?
Ha egy sejt teljes DNS-dupla spirálját, azaz körülbelül 3,4x109 bázispárt összekapcsolnák, ez több mint egy méter hosszúságot eredményezne. Az összes hozzáadott kromoszóma teljes hossza csak körülbelül 115 µm. Ezt a hosszúságbeli különbséget a kromoszómák nagyon kompakt szerkezete magyarázza, amelyben a DNS-t nagyon specifikus módon többször feltekerik vagy spirálba helyezik.
Ebben fontos szerepet játszanak a hisztonok, a fehérjék speciális formája. Összesen 5 különböző hiszton létezik: H1, H2A, H2B, H3 és H4. Az utolsó négy hiszton közül kettő egy hengeres szerkezetet alkot, az oktámert, amely körül a kettős spirál körülbelül kétszer kanyarodik (= szuper spirál). A H1 stabilizálása érdekében ehhez a szerkezethez kapcsolódik.
Ezt a DNS, oktamer és H1 komplexet nukleoszómának nevezzük. Ezen nukleoszómák közül több olyan, mint egy gyöngysor, viszonylag rövid időközönként (10-60 bázispár) egymás mögött. A kromoszómák közötti szakaszokat spacer DNS-nek nevezzük. Az egyes nukleoszómák most újra érintkezésbe kerülnek a H1-en keresztül, ami további spirálozást és ezáltal kompressziót is létrehoz.
A kapott szál viszont olyan hurkokban van, amelyeket stabil savas, nem hiszton fehérjékből álló gerinc stabilizál, más néven hertonok. Ezek a hurkok viszont a fehérjék által stabilizált spirálokban vannak jelen, ami a kompresszió utolsó szakaszát eredményezi. Ez a nagyfokú kompresszió azonban csak a mitózis alatti sejtosztódás összefüggésében fordul elő.
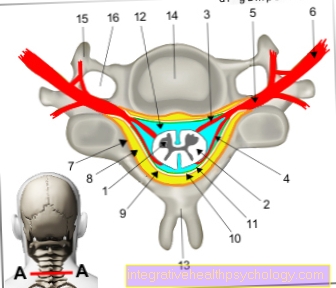
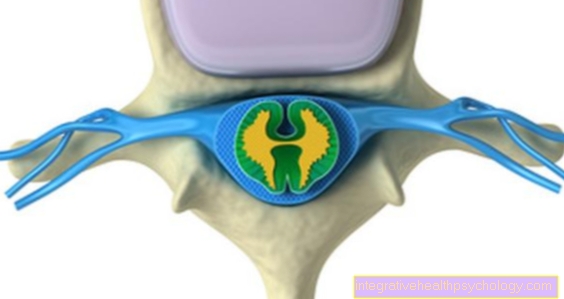
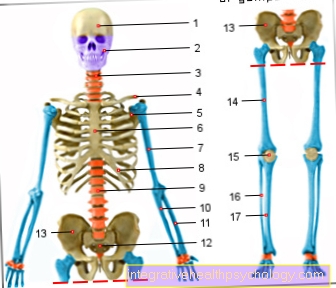
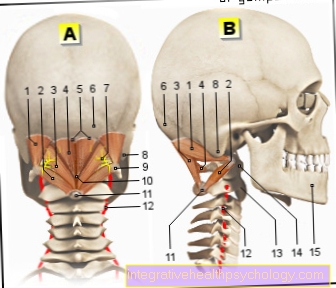
Ebben a fázisban a kromoszómák jellegzetes alakja is látható, amely két kromatidából áll. A helyet, ahol ezek összekapcsolódnak, centromernek nevezzük. Minden metafázisos kromoszómát két rövid és két hosszú karra oszt, amelyeket p és q karoknak is neveznek.
Ha a centromér nagyjából a kromoszóma közepén fekszik, akkor metacentrikus kromoszómának nevezzük, ha teljes egészében az akrocentrikus kromoszóma egyik végén fekszik. A köztük lévőket szubmetacentrikus kromoszómáknak nevezzük. Ezek a fénymikroszkóp alatt már látható különbségek a hosszúsággal együtt lehetővé teszik a kromoszómák kezdeti osztályozását.
Mik a telomerek?
A telomerek a kromoszómák végei ismétlődő szekvenciákkal (TTAGGG). Ezek nem tartalmaznak releváns információt, inkább a relevánsabb DNS-szakaszok elvesztésének megakadályozására szolgálnak. Minden sejtosztódással a kromoszóma egy része elvész a DNS-replikáció mechanizmusa révén.
Tehát a telomerek bizonyos értelemben pufferek, amelyek késleltetik azt a pontot, amikor a sejt elválik fontos információtól. Ha egy sejt telomerje kevesebb, mint 4000 bázispár hosszú, elindul a programozott sejthalál (apoptózis). Ez megakadályozza a hibás genetikai anyag terjedését a szervezetben. Néhány sejtben vannak telomerázok, azaz enzimek, amelyek képesek ismét meghosszabbítani a telomereket.
Az őssejteken kívül, amelyekből az összes többi sejt származik, ezek csírasejtek és az immunrendszer bizonyos sejtjei. Ezenkívül telomerázok találhatók a rákos sejtekben is, ezért beszélhetünk egy sejt megörökítéséről ebben az összefüggésben.
Itt olvashat el mindent a témáról: Telomerek - anatómia, funkció és betegségek
Mi az a kromatin?
A kromatin a sejtmag teljes tartalmára utal, amelyet bázissal lehet festeni. Ezért a DNS mellett a kifejezés bizonyos fehérjéket is tartalmaz, például hisztonokat és hertonokat (lásd a szerkezetet), valamint bizonyos RNS-fragmenseket (hn és snRNS).
A sejtciklus fázisától vagy a genetikai aktivitástól függően ez az anyag különböző sűrűségben áll rendelkezésre. A sűrűbb formát heterokromatinnak nevezzük. A könnyebb megértés érdekében tehát „tárolási formának” tekinthetjük, és itt ismét különbséget tehetünk konstitutív és fakultatív heterokromatin között.
A konstitutív heterokromatin a legsűrűbb forma, amely a sejtciklus minden fázisában a legmagasabb kondenzációban van jelen. Ez az emberi genom körülbelül 6,5% -át teszi ki, és főként a centromerek és a kromoszóma karok (telomerek) végei közelében található kis mértékben, de más helyeken is (elsősorban az 1., 9., 16., 19. és Y. Kromoszóma) . Ezenkívül a konstitutív heterokromatin nagy része a magmembrán közelében, vagyis a sejtmag szélén helyezkedik el. A közepén lévő helyet az aktív kromatinnak, az euchromatinnak tartják fenn.
A fakultatív heterokromatin kissé kevésbé sűrű, aktiválható és deaktiválható szükség szerint vagy a fejlődési stádiumtól függően. Jó példa erre a női kariotípusok második X-kromoszómája. Mivel egy X kromoszóma alapvetően elegendő a sejt életben maradásához, mint ami végső soron a férfiak számára is elegendő, a kettő egyike inaktiválódik az embrionális fázisban. a dezaktivált X kromoszóma Barr test néven ismert.
Csak a sejtosztódás során, a mitózis összefüggésében kondenzálódik teljesen, ezáltal a metafázisban éri el a legnagyobb kompressziót. Mivel azonban a különböző géneket gyakran másképp olvassák - elvégre nem minden fehérjére van szükség mindig azonos mennyiségben -, itt is különbséget tesznek az aktív és az inaktív euchromatin között.
További információ erről:: Chromatin
Haploid kromoszómák
A haploid (görög haploos = egyedülálló) azt jelenti, hogy a sejt összes kromoszómája külön-külön van jelen, vagyis nem párban (diploid), mint általában. Ez az összes petesejt és spermium sejt természetes állapota, amelyben a két egyforma kromatidát az első meiózis során kezdetben nem választják el, hanem ehelyett az összes kromoszómapárt különválasztják.
Ennek eredményeként az első meiózis után az emberek lánysejtjeiben a szokásos 46 kromoszóma helyett csak 23 van, ami a kromoszóma haploid halmazának felének felel meg. Mivel ezeknek a leánysejteknek még mindig mindegyik 2 kromoszómából álló kromoszómájának azonos másolata van, szükség van a második meiózisra, amelyben a két kromatid el van választva egymástól.
Politén kromoszómák
A politén kromoszóma olyan kromoszóma, amely nagyszámú, genetikailag azonos kromatidból áll. Mivel az ilyen kromoszómák még nagyítás esetén is könnyen láthatók, néha óriás kromoszómának nevezik őket. Ennek előfeltétele az endoreplikáció, amelynek során a sejtmag kromoszómáit többször megsokszorozzák anélkül, hogy sejtosztódás történne.
Melyek a kromoszómák funkciói?
A kromoszóma, mint genomunk szervezeti egysége, elsősorban annak biztosítását szolgálja, hogy a megduplázódott genom a sejtosztódás során egyenletesen oszlik meg a leánysejtek között. Ehhez érdemes közelebbről megvizsgálni a sejtosztódás mechanizmusait vagy a sejtciklust:
A sejt a ciklus nagy részét az interfázisban tölti, ami azt az egész időtartamot jelenti, amelyben a sejt nem azonnal készül megosztani. Ez viszont G1, S és G2 fázisokra oszlik.
A G1 fázis (G, mint rés, azaz rés) azonnal követi a sejtosztódást. Itt a sejt ismét megnövekszik, és általános metabolikus funkciókat lát el.
Innen átállhat a G0 fázisra is. Ez azt jelenti, hogy olyan szakaszra vált, amely már nem képes megosztódni, és normális esetekben nagymértékben megváltozik egy nagyon specifikus funkció (sejtdifferenciálás) teljesítése érdekében is. E feladatok teljesítése érdekében a nagyon specifikus géneket intenzívebben, másokat kevésbé vagy egyáltalán nem olvassuk.
Ha a DNS-szegmensre sokáig nincs szükség, gyakran a kromoszómák azon részein helyezkedik el, amelyek hosszú ideig sűrűn vannak tele (lásd a kromatint). Ennek egyrészt a helytakarékosság a célja, de a génszabályozás egyéb mechanizmusai mellett további védelmet nyújt a véletlenszerű elolvasás ellen is. Megfigyelték azonban azt is, hogy nagyon specifikus körülmények között a G0 fázisból differenciált sejtek visszatérhetnek a ciklusba.
A G1 fázist követi az S fázis, vagyis az a fázis, amelyben új DNS szintetizálódik (DNS replikáció). Itt az egész DNS-nek a leglazább formájában kell lennie, vagyis az összes kromoszóma teljesen tekercseletlen (lásd a szerkezetet).
A szintézis fázis végén a teljes genetikai anyag két példányban van jelen a sejtben. Mivel a kópia a centromérán keresztül még mindig az eredeti kromoszómához kapcsolódik (lásd a szerkezetet), nem beszélünk a kromoszómák duplikációjáról.
Minden kromoszóma most egy helyett két kromatidból áll, így később a mitózis során felveheti a jellegzetes X-alakot (szigorúan véve az X-forma csak a metacentrikus kromoszómákra vonatkozik). A következő G2 fázisban az azonnali felkészülés zajlik a sejtosztódásra. Ez magában foglalja a replikációs hibák és a szálszakadások részletes ellenőrzését is, amelyeket szükség esetén meg lehet javítani.
Alapvetően kétféle sejtosztódás létezik: a mitózis és a meiózis. A csírasejtek kivételével egy szervezet összes sejtje mitózis révén keletkezik, amelynek egyetlen feladata két genetikailag azonos leánysejt kialakulása.
A meiózis célja viszont genetikailag különböző sejtek létrehozása:
Első lépésben a megfelelő (homológ), de nem azonos kromoszómákat felosztjuk. Csak a következő lépésben választják el a két azonos kromatidból álló kromoszómákat, és ismét elosztják két-két leánysejtben, így végül egy prekurzor sejtből négy különböző genetikai anyagú csírasejt keletkezik.
A kromoszómák formája és felépítése elengedhetetlen mindkét mechanizmushoz: Speciális "fehérjefonalak", az úgynevezett orsó készülék kapcsolódnak az erősen kondenzált kromoszómákhoz, és finoman szabályozott folyamatban húzzák a kromoszómákat a középsíkról (egyenlítői síkról). az egy körüli sejt ellentétes pólusaira az egyenletes eloszlás biztosítása érdekében. A kromoszómák mikrostruktúrájának apró változásai is itt súlyos következményekkel járhatnak.
Minden emlősnél az X és Y nemi kromoszómák aránya meghatározza az utódok nemét is. Alapvetően minden attól függ, hogy a petesejttel egyesülő spermiumnak van-e X vagy Y kromoszómája. Mivel a spermium mindkét formája mindig pontosan azonos mértékben termelődik, a valószínűség mindkét nem esetében mindig kiegyensúlyozott. Ez a véletlenszerű rendszer egyenletesebb nemi eloszlást garantál, mint például a környezeti tényezők, például a hőmérséklet esetén.
Tudjon meg többet a témáról: Sejtmag osztódás
Hogyan adódik tovább a genetikai felépítés a kromoszómákon keresztül?
Ma már tudjuk, hogy a tulajdonságok olyan gének révén öröklődnek, amelyeket a sejtek DNS formájában tárolnak. Ezek viszont 46 kromoszómára oszlanak, amelyeken a 25 000-30000 emberi gén oszlik meg.
Magán a fenotípusnak nevezett tulajdonság mellett létezik genetikai megfelelője is, amelyet genotípusnak neveznek. Azt a helyet, ahol egy gén a kromoszómán található, lókusznak nevezzük. Mivel az embereknek minden kromoszómájuk duplája, minden gén kétszer is előfordul. Az egyetlen kivétel ez alól a férfiak X-kromoszóma-génje, mivel az Y-kromoszóma csak az X-kromoszómán található genetikai információ töredékét hordozza magában.
Különböző géneket, amelyek ugyanazon a lokuszon vannak, alléloknak nevezzük. Gyakran több mint két különböző allél található egy lokuszban. Ezután a polimorfizmusról beszélünk. Egy ilyen allél egyszerűen lehet ártalmatlan variáns (normális variáns), de akár kóros mutációk is, amelyek kiválthatják az örökletes betegséget.
Ha egyetlen gén mutációja elegendő a fenotípus megváltoztatásához, akkor monogén vagy Mendeli öröklésről beszélhetünk. Az örökölhető tulajdonságok közül sok azonban több kölcsönhatásban lévő gén révén öröklődik, ezért sokkal nehezebb tanulmányozni.
Mivel az anya és az apa mindkét génjük egyikét adja át a gyermeknek egy mendeli örökségben, a következő generációban mindig négy lehetséges kombináció létezik, amelyek ezek egy tulajdonság vonatkozásában is azonosak lehetnek. Ha az egyén mindkét allélja ugyanolyan hatást gyakorol a fenotípusra, akkor az egyén e jellemző tekintetében homozigóta, és a jellemző ennek megfelelően teljes mértékben kifejeződik.
A heterozigótáknak két különböző allélja van, amelyek különböző módon képesek kölcsönhatásba lépni egymással: Ha az egyik allél domináns a másik felett, akkor teljesen elnyomja az expresszióját, és a domináns tulajdonság láthatóvá válik a fenotípusban. Az elnyomott allélt recesszívnek nevezzük.
Kodomináns öröklés esetén mindkét allél kifejezheti magát, és nem befolyásolja egymást, míg a köztes öröklésnél mindkét jellemző keveredik. Jó példa erre az AB0 vércsoport-rendszer, amelyben A és B együtt domináns egymással, de 0 dominál egymással szemben.
Mi a normális kromoszóma halmaz az emberben?
Az emberi sejtekben 22 nemtől független kromoszómapár (autoszómák) és két nemi kromoszóma (gonoszóma) van, így összesen 46 kromoszóma alkotja a kromoszómák egyik készletét.
Az autoszómák általában párban érkeznek. Egy pár kromoszómája hasonló alakú és génszekvenciájú, ezért homológnak nevezik őket. A nők két X kromoszómája szintén homológ, míg a férfiak X és Y kromoszómával rendelkeznek. Ezek oly módon különböznek a jelenlévő gének formájától és számától, hogy a homológiáról már nem lehet beszélni.
A csírasejtekben, azaz a petesejtekben és a spermium sejtekben csak a kromoszóma fele van a meiózis miatt, nevezetesen 22 egyedi autoszóma és egy-egy gonoszóma. Mivel a megtermékenyítés során a csírasejtek összeolvadnak, és néha teljes szegmenseket cserélnek (keresztezés), létrejön a kromoszómák új kombinációja (rekombináció). Valamennyi kromoszómát kariotípusnak nevezzük, amely néhány kivételtől eltekintve (lásd a kromoszóma-rendellenességeket) azonos neműek mindegyikében azonos.
Itt mindent megtudhat a témáról: Mitózis - egyszerűen magyarázható!
Miért vannak mindig kromoszómapárok?
Alapvetően erre a kérdésre egy mondattal lehet megválaszolni: Mert hasznosnak bizonyult.A kromoszómapárok jelenléte és a rekombináció elve elengedhetetlen az öröklődés szempontjából a nemi szaporodás szempontjából. Ily módon egy teljesen új egyén véletlenül előkerülhet két egyén genetikai anyagából.
Ez a rendszer hatalmas mértékben növeli a fajok tulajdonságainak változatosságát, és biztosítja, hogy sokkal gyorsabban és rugalmasabban tudjon alkalmazkodni a megváltozott környezeti viszonyokhoz, mint azt csak mutáció és szelekció révén lehetne megvalósítani.
A kettős kromoszómasorozatnak védő hatása is van: ha egy gén mutációja a funkció meghibásodásához vezetne, akkor is van egyfajta "biztonsági másolat" a második kromoszómában. Ez nem mindig elegendő ahhoz, hogy a szervezet kompenzálja a meghibásodást, különösen, ha a mutált allél domináns, de növeli annak esélyét. Ráadásul így a mutáció nem jut tovább automatikusan minden utódra, ami viszont megvédi a fajt a túlságosan radikális mutációktól.
Mi a kromoszóma mutáció?
Genetikai hibák származhatnak ionizáló sugárzásból (pl. Röntgen), vegyi anyagokból (pl. A cigarettafüstben lévő benzopirolból), bizonyos vírusokból (pl. HP vírusok), vagy kis valószínűséggel pusztán véletlenül is felmerülhetnek. Gyakran több tényező is szerepet játszik a fejlődésében. Elvileg ilyen változások a test összes szövetében előfordulhatnak, de gyakorlati okokból az elemzés általában a limfocitákra (az immunsejtek speciális típusa), a fibroblasztokra (kötőszöveti sejtek) és a csontvelő sejtekre korlátozódik.
A kromoszóma mutáció az egyes kromoszómák egyik fő szerkezeti változása. A teljes kromoszómák hiánya vagy hozzáadása viszont genom vagy ploidia mutáció lenne, míg a génmutáció kifejezés viszonylag kicsi változásokat jelent egy génen belül. A kromoszóma-aberráció (latin aberrare = eltérni) kifejezés valamivel tágabb és magában foglal minden, a fénymikroszkóppal kimutatható változást.
A mutációknak nagyon különböző hatása lehet:
- A csendes mutációk, vagyis azok a mutációk, amelyekben a változásnak nincs hatása az egyénre vagy utódaikra, meglehetősen atipikusak a kromoszóma-rendellenességekre, és gyakrabban fordulnak elő a gén- vagy pontmutációk területén.
- Az egyik a funkcióvesztés mutációjáról beszél, amikor a mutáció rosszul összehajtott és ezért működőképtelen fehérjét eredményez, vagy egyáltalán nem tartalmaz fehérjét.
- Az úgynevezett funkciógyarapodási mutációk megváltoztatják a hatás típusát vagy a termelt fehérjék mennyiségét oly módon, hogy teljesen új hatások jelentkeznek. Ez egyrészt döntő mechanizmus az evolúció és így egy faj fennmaradása vagy új fajok megjelenése szempontjából, de másrészt, mint a Philadelphia kromoszóma esetében, ez is döntő mértékben hozzájárulhat a a rákos sejtek fejlődése.
A kromoszóma-rendellenességek különböző formái közül a legismertebbek valószínűleg a numerikus eltérések, amelyekben az egyes kromoszómák csak egyszer (monoszómia) vagy akár háromszorosak (trisomia) vannak jelen.
Ha ez csak egyetlen kromoszómára vonatkozik, akkor azt aneuploidiának nevezzük, és a teljes kromoszóma készletet a poliploidia (tri- és tetraploidia) befolyásolja. A legtöbb esetben ez a maldisztribúció a csírasejt-fejlődés során keletkezik a kromoszómák szétválasztása (nem diszjunkciója) révén a sejtosztódás (meiózis) során. Ez a kromoszómák egyenetlen eloszlásához vezet a leánysejteken, és ezáltal a gyermek számszerű aberrációjához vezet.
A nem nemi kromoszómák (= autoszómák) monoszómiái nem kompatibilisek az élettel, ezért élő gyermekeknél nem fordulnak elő. A 13., 18. és 21. triszómiák kivételével az autoszomális triszómiák szinte mindig spontán abortuszokhoz vezetnek.
Mindenesetre, a nemi kromoszómák aberrációival ellentétben, amelyek szintén nem feltűnőek, mindig vannak súlyos klinikai tünetek, és általában többé-kevésbé kifejezett külső rendellenességek (diszmorfizmusok).
Ilyen maldistribúció az élet későbbi szakaszaiban is előfordulhat mitotikus sejtosztódással (a csírasejtek kivételével minden sejt). Mivel az érintett sejteken kívül változatlan sejtek vannak, szomatikus mozaikról beszélünk. A szomatikus (görög soma = test) kifejezés alatt minden olyan sejtet értünk, amely nem csírasejt. Mivel a test sejtjeinek csak egy kis része érintett, a tünetek általában sokkal enyhébbek. Ezért a mozaiktípusokat sokáig sokáig nem fedezik fel.
Itt mindent megtudhat a témáról: Kromoszóma mutáció
Mi a kromoszóma-rendellenesség?
A strukturális kromoszóma-rendellenesség alapvetően megfelel a kromoszóma mutáció definíciójának (lásd fent). Ha a genetikai anyag mennyisége változatlan marad, és egyszerűen másképp oszlik el, akkor kiegyensúlyozott aberrációról beszélünk.
Ez gyakran transzlokáció útján történik, vagyis egy kromoszómaszegmens átvitele egy másik kromoszómába. Ha ez két kromoszóma közötti csere, akkor az egyik kölcsönös transzlokációról beszél. Mivel a fehérjék termeléséhez a genomnak csak körülbelül 2% -ára van szükség, nagyon kicsi a valószínűsége annak, hogy egy ilyen gén a törésponton van, és ezáltal elveszíti funkcióját vagy károsodik benne. Ezért egy ilyen kiegyensúlyozott aberráció gyakran észrevétlen marad, és több generáción keresztül továbbadódik.
Ez azonban a csírasejtek fejlődése során a kromoszómák rossz eloszlásához vezethet, amely meddőséghez, spontán vetélésekhez vagy kiegyensúlyozatlan aberrációjú utódokhoz vezethet.
A kiegyensúlyozatlan aberráció spontán is előfordulhat, vagyis családtörténet nélkül. Annak a valószínűsége, hogy a gyermek életben, kiegyensúlyozatlan aberrációval születik, nagymértékben függ az érintett kromoszómáktól, és 0 és 60% között változik. Ez egy kromoszómaszegmens elvesztéséhez (= törlés) vagy megkettőzéséhez (= duplikációjához) vezet. Ebben az összefüggésben a részleges mono- és triszómiákról is beszélünk.
Bizonyos esetekben ezek két különböző régióban fordulnak elő együtt, a részleges monoszómia általában meghatározóbb a klinikai tünetek előfordulásában. Ezek a törlés kiemelkedő példái Cat Scream szindróma és Wolf-Hirschhorn szindróma.
Mikrodeletióról akkor beszélünk, amikor a változás már nem határozható meg a fénymikroszkóppal, vagyis amikor egy vagy néhány gén elvesztéséről van szó. Ezt a jelenséget tekintik a Prader-Willi-szindróma és az Angelman-szindróma okának, és szorosan összefügg a retionoblastoma kialakulásával.
A Robertson-áthelyezés különleges eset:
Két akrocentrikus kromoszóma (13, 14, 15, 21, 22) centromerájukon egyesülve egyetlen kromoszómát képeznek, miután a rövid karok elvesznek (lásd a szerkezetet). Bár ez kromoszómák számának csökkenését eredményezi, ezt kiegyensúlyozott aberrációnak nevezik, mivel ezekben a kromoszómákban a rövid karok elvesztése könnyen kompenzálható. Itt is gyakran csak a következő generációkban észlelhetők a hatások, mivel nagyon nagy a valószínűsége a vetéléseknek vagy a triszómiában élő gyermekeknek.
Ha egy kromoszómán belül két törés van, akkor előfordulhat, hogy a köztes szegmens 180 ° -kal elfordul és beépül a kromoszómába. Ez az inverziónak nevezett folyamat csak akkor kiegyensúlyozatlan, ha a töréspont egy aktív génben található (a teljes genetikai anyag 2% -a). Attól függően, hogy a centromér az invertált szegmensen belül vagy kívül helyezkedik el, peri- vagy paracentrikus inverzióról van szó. Ezek a változások hozzájárulhatnak a genetikai anyag egyenetlen eloszlásához is a csírasejteken.
Paracentrikus inverzióban, amelyben a centroméra nincs az invertált szegmensben, két vagy egyáltalán nem centromérás csírasejtek is megjelenhetnek. Ennek eredményeként a megfelelő kromoszóma elvész a legelső sejtosztódások során, ami szinte biztosan vetéléshez vezet.
Az inszerció egy kromoszómatöredék máshova történő telepítése. Itt is elsősorban az utódokat érinti hasonló módon. Különösen a végdarabok törlése után fordulhat elő gyűrűs kromoszóma. A szekvenciák típusa és mérete meghatározó a tünetek súlyossága szempontjából. Ez ráadásul helytelen eloszlásokhoz, és ezáltal mozaiktípusokhoz vezethet a testsejteken belül.
Ha a metafázisos kromoszóma helytelenül elválik a sejtosztódás során, akkor izokromoszómák keletkezhetnek. Ez két pontosan ugyanaz a kromoszóma, amely csak hosszú vagy csak rövid karokból áll. Az X kromoszóma esetében ez Ulrich-Turner szindrómaként (X monoszómia) nyilvánulhat meg.
További információ erről a témáról: Kromoszóma-rendellenesség
Trisomy 21
A 21. triszómia, közismertebb nevén Down-szindróma, valószínűleg a leggyakoribb numerikus kromoszóma-rendellenesség az élő születések között, a hímeket kissé gyakrabban érintik (1,3: 1).
A 21-es triszómia bekövetkezésének valószínűsége különféle demográfiai tényezőktől függ, például az anyák átlagos születési életkorától, és régiónként kissé változó.
A 21-es triszómia 95% -a a meiózis (csírasejt-osztódás) összefüggésében bekövetkező osztódási hiba, nevezetesen a nem diszjunkció, vagyis a testvérkromatidok elválasztásának sikertelensége miatt keletkezik.
Ezeket szabad triszómiáknak nevezik, és 90% -ban az anyai, 5% az apai és további 5% az embrionális genomban fordulnak elő.
További 3% a 14-es vagy 21-es kromoszómán lévő kiegyensúlyozatlan transzlokációkból származik; 21 transzlokáció, létrehozva egy normális és egy kettős kromoszómát 21. A fennmaradó 2% olyan mozaiktípus, amelyben a triszómia nem a csírasejtekben következett be, ezért nem érinti az összes testsejtet. A mozaiktípusok gyakran annyira enyheek, hogy hosszú ideig teljesen észrevétlenek maradhatnak.
Mindenesetre kromoszóma vizsgálatot kell végezni annak érdekében, hogy megkülönböztessük a tünetekkel azonos azonos szabad triszómiát az esetleg öröklődő transzlokációs triszómiától. Ezután következhet az előző generációk családtörténete.
Érdekel ez a téma? Olvassa el a következő cikket erről: Trisomy 21
13. triszómia
A Trisomy 13 vagy a Patau szindróma gyakorisága 1: 5000, és sokkal ritkább, mint a Down szindróma. Az okok (szabad triszómiák, transzlokációk és mozaiktípusok) és százalékos eloszlásuk nagyrészt azonos.
Elméletileg szinte minden eset prenatálisan diagnosztizálható ultrahang vagy PAPP-A teszt segítségével. Mivel a PAPP-A teszt nem feltétlenül része a rutinvizsgálatoknak, Közép-Európában az esetek körülbelül 80% -át születés előtt diagnosztizálják.
Az ultrahangon már látható növekedési maradvány, kétoldali ajak- és szájpadhasadék, szokatlanul kicsi szemek (microphthalmia). Ezenkívül általában az előagy és az arc különböző súlyosságú rendellenességei vannak (holoprosencephaly).
Míg a lobár formában az agyféltekék szinte teljesen el vannak választva, és laterális kamrák jönnek létre, addig a féllobár formában gyakran csak az agy hátsó része van elválasztva, és az oldalsó kamrák hiányoznak. A legsúlyosabb formában, az alobár formában nincs elválasztva az agyféltekék.
A fél- vagy alabár alakú csecsemők általában születésük után azonnal meghalnak. Egy hónap elteltével a halálozási arány az élveszületések 50% -a. 5 éves korig a 13. triszómia halálozási aránya 90% -ra emelkedik. Az agy rendellenességei miatt a legtöbb esetben a betegek egész életen át ágyhoz vannak kötve, és nem tudnak beszélni, ezért a teljes ellátástól függenek. Emellett a Trismoie 13 messzemenő fizikai megnyilvánulásai is lehetnek.
További információ a témáról: Trisomy 13 a születendő gyermeknél
Trisomy 16
Alapvetően a triszómia 16 a leggyakoribb (az összes triszómia körülbelül 32% -a), de a 16-os triszómiában élő gyermekek nagyon ritkák. Az élveszületések általában csak részleges triszómiákban vagy mozaiktípusokban fordulnak elő. A triszómiák közül azonban a leggyakrabban felelős a halandó születésekért: a kromoszóma-rendellenességek miatti 100 vetélésből 32-re vezethető vissza a trisomia ezen formája.
Ezért főleg prenatális, azaz prenatális, azonosítható jellemzőket dokumentáltak. Figyelemre méltó itt a különféle szívhibák, a növekedés lelassulása, egyetlen köldökartéria (egyébként kettős) és a nyak megnövekedett átlátszósága, ami a még nem teljesen kifejlődött nyirokrendszer miatti folyadékfelhalmozódással és a bőr ezen a területen megnövekedett rugalmasságával magyarázható. Ezenkívül a fiziológiás köldöksérv, vagyis a bél nagy részének a köldökön keresztüli ideiglenes elmozdulása kifelé gyakran nem regresszálódik megfelelően, amelyet omphalocele vagy köldökzsinórtörésnek neveznek.
Keresztezett ujjakkal ellátott hajlító kontraktúra ultrahangon is gyakran kimutatható. A kevés élveszületésben észrevehető az általános izom hipotenzió, vagyis az általános izomgyengeség. Ez ivásgyengeséghez vezet, és biztosíthatja, hogy a csecsemőt mesterségesen kell táplálni. Gyakran előfordul a négyujjas barázda is, amely annyira jellemző a triszómiákra. Itt is a trisomia előfordulásának gyakorisága közvetlenül összefügg az anya életkorával.
Trisomy 18
Az Edwards-szindróma, azaz a 18-as triszómia 1: 3000 gyakorisággal fordul elő. A prenatális diagnosztikával megegyezik a Patau-szindrómával: Ugyanezen vizsgálatok itt is lehetővé tennék, hogy az összes beteget teljesen megtalálják a születés előtt. Az okok és eloszlásuk összehasonlítható más triszómiákkal (lásd a 21. triszómiát).
Ezenkívül a 18. triszómiában részleges triszómiák fordulnak elő, amelyek a mozaiktípusokhoz hasonlóan sokkal enyhébb klinikai lefolyásokhoz vezetnek. A kapcsolódó diszmorfizmusok az Edwards-szindrómára is rendkívül jellemzőek: Születéskor a betegek testtömege nagymértékben csökken, 2 kg (normál: 2,8–4,2 kg), a visszahúzódó széles homlok, az arc általában fejletlen alsó fele, kis szájjal nyíló, keskeny szemhéj rések és hátrafelé fordított, alakváltozott fülek (faun fül). Az újszülött számára szokatlanul erősen fejlett fej hátsó része is észrevehető. A bordák szokatlanul keskenyek és törékenyek. Az újszülötteknél az egész izomzat is állandó feszültséggel rendelkezik (tónus), amely azonban a túlélőkben az első hetek után visszafejlődik.
Egy másik jellemző jellemző a 2. és az 5. ujj keresztezése a 3. és 4. felett, az összes ujj beütésével, míg a lábak szokatlanul hosszúak (elteltek), különösen hangsúlyos sarokkal, elakadt lábujjakkal és hátratett nagy lábujjal rendelkeznek. .
A súlyos szervi rendellenességek gyakoriak és általában együttesen fordulnak elő: szív- és veseelégtelenségek, a bél rosszul összecsapódása (malrotációja), a hashártya (mesenterium commune) tapadásai, a nyelőcső elzáródása (nyelőcső-atresia) és még sok más.
Ezen malformációk miatt a halálozási arány az első 4 napban körülbelül 50%, csak körülbelül 5-10% él egy évesnél idősebb korban. Az abszolút kivétel a felnőttkorban való túlélés, mindenesetre az értelmi fogyatékosság nagyon hangsúlyos, nem tud beszélni, ágyhoz kötött és inkontinens, tehát teljesen függ a külső segítségtől.
A 18. triszómiával kapcsolatos további információkért kérjük, olvassa el a témáról szóló részletes cikkünket is:
- Trisomy 18 (Edwards-szindróma)
- 18. triszómia a születendő gyermeknél
Trisomy X
Az X triszómia a numerikus kromoszóma-rendellenesség legszembetűnőbb formája, az érintettek logikusan mind nőstény megjelenése nem különbözik nagyban a többi nőtől. Néhányan kiemelkednek, mert különösen magasak és kissé „kövér” arcvonásokkal rendelkeznek. A mentális fejlődés szintén nagyrészt normális lehet, a határértéktől az enyhe értelmi fogyatékosságig terjed.
Ez az intelligenciahiány azonban valamivel súlyosabb, mint a nemi kromoszómák többi triszómiájával (XXY és XYY). 1: 1000 gyakorisággal valójában nem olyan ritka, de mivel a triszómia általában nem jár klinikailag jelentős tünetekkel, a betegségben szenvedő nők többségét valószínűleg soha nem diagnosztizálják egész életében.
A hordozókat többnyire véletlenül fedezik fel családi ellenőrzés vagy a prenatális diagnosztika során.A termékenység kissé csökkenthető, és a nemi kromoszóma-rendellenességek aránya a következő generációban kissé megnőhet, ezért genetikai tanácsadás javasolt, ha gyermekeket szeretne.
Csakúgy, mint a többi triszómiánál, az X-triszómia is leggyakrabban szabad triszómiaként alakul ki, vagyis a testvérkromatidák osztódásának (nem disszjunkciójának) hiánya miatt. Itt is általában az anyai petesejtek érése során merül fel, bár a valószínűség az életkor előrehaladtával növekszik.
Törékeny X szindróma
A törékeny X-szindrómát vagy a Martin Bell-szindrómát előnyben részesítik a férfiaknál, mivel csak egy X-kromoszómájuk van, ezért őket jobban érinti a változás.
1: 1250 gyakorisággal fordul elő az élő férfi születések között egy év alatt, ezáltal a nem specifikus mentális retardáció leggyakoribb formája, vagyis minden olyan mentális fogyatékosság, amelyet nem lehet jellemző tünetekkel járó speciális szindrómával leírni.
A törékeny X szindróma a lányoknál általában valamivel gyengébb formában fordulhat elő, ami az egyik X kromoszóma véletlen inaktiválásának köszönhető. Minél nagyobb a kikapcsolt egészséges X kromoszóma aránya, annál erősebbek a tünetek.
Legtöbbször azonban a nők hordozzák az premutációt, amely még nem okoz klinikai tüneteket, de masszívan növeli a teljes mutáció valószínűségét fiaikban. Nagyon ritka esetekben a férfiak is hordozhatják a premutációt, amelyet aztán csak olyan lányoknak adhatnak át, akik azonban általában klinikailag egészségesek (Sherman-paradoxon).
A szindrómát az FMR-génben (egy törzshely-mentális retardáció) rendkívül megnövekedett számú CGG-hármas váltja ki (egy bizonyos bázisszekvencia); 10-50 kópia helyett az 50-200 premutáció, amikor teljesen kifejlődött 2000 példány.
A fénymikroszkóp alatt ez úgy néz ki, mint egy törés a hosszú karban, ez adta a szindróma nevét. Ez az érintett gén dezaktiválásához vezet, ami viszont a tüneteket okozza.
Az érintett emberek lassú beszéd- és mozgásfejlődést mutatnak, és viselkedési problémákat mutathatnak, amelyek a hiperaktivitás, de az autizmus irányába is elmozdulhatnak. A pusztán külső rendellenességek (a diszmorfizmus jelei) egy hosszú arc, kiemelkedő állal és kiálló fülekkel. A pubertáskor a herék gyakran nagymértékben megnagyobbodnak (makrosorchidia), és az arcvonások durvábbá válnak. A premutáció női hordozói között enyhén halmozódik a pszichés rendellenességek és egy különösen korai menopauza.
Mi a kromoszóma-elemzés?
A kromoszóma-elemzés a citogenetika folyamata, amelynek során numerikus vagy strukturális kromoszóma-rendellenességek mutathatók ki.
Ilyen elemzést alkalmaznának például abban az esetben, ha kromoszóma-szindróma gyanúja merül fel, azaz fejlődési rendellenességek (diszmorfizmusok) vagy értelmi fogyatékosságok (retardáció) esetén, de meddőség, rendszeres vetélések (abortuszok) esetén is, bizonyos rákos megbetegedések (pl. limfómák vagy leukémia).
Ehhez általában limfocitákra van szükség, egy speciális típusú immunsejtre, amelyet a beteg véréből nyernek. Mivel ily módon csak viszonylag kis mennyiség nyerhető, a sejteket stimuláljuk a fitohemagglutininnel való osztódásra, majd a limfocitákat a laboratóriumban tenyészthetjük.
Bizonyos esetekben mintákat (biopsziákat) vesznek a bőrből vagy a gerincvelőből, hasonló eljárással. A cél a lehető legtöbb DNS-anyag megszerzése, amely jelenleg a sejtosztódás közepén van. A metafázisban az összes kromoszóma egy szinten helyezkedik el, megközelítőleg a sejt közepén, annak érdekében, hogy a következő lépésben, az anafázisban a sejt ellentétes oldalaihoz (pólusaihoz) vonzódjanak.
Ebben az időpontban a kromoszómák különösen szorosan vannak csomagolva (erősen kondenzálódnak). Az orsóméreg kolchicint adunk hozzá, amely pontosan a sejtciklus ezen szakaszában működik, így a metafázisos kromoszómák felhalmozódnak. Ezután speciális festési módszerekkel izolálják és megfestik őket.
A leggyakoribb a GTG sávosodás, amelynek során a kromoszómákat tripszinnel, emésztőenzimmel és Giemsa pigmenttel kezelik. A különösen sűrűn tömött, valamint az adeninben és a timinnel gazdag területek sötétek.
Az így kapott G-sávok jellemzőek az egyes kromoszómákra, és leegyszerűsítve a kevesebb gént tartalmazó régióknak tekinthetők. Az így festett kromoszómákról ezerszeres nagyítással készül a kép, és egy számítógépes program segítségével kariogram készül. A sávminta mellett a kromoszóma méretét és a centromér helyzetét is felhasználják a kromoszómák ennek megfelelő elrendezésében. De vannak más sávolási módszerek is, amelyeknek nagyon különböző előnyei lehetnek.
A szerkesztőség ajánlásai
Általánosabb információkért lásd a következő cikkeket:
- Sejtmag osztódás
- A sejtmag funkciói
- Trisomy 21
- Genetikai betegségek